Transforming Human Health
Driving discovery, advancing care and educating leaders

Our Schools
Advancing Care
News
- April 2, 2025
A new AI tool that uses machine learning and nurses' instincts detects patient deterioration nearly two days faster than conventional methods, reducing risk of death and complications.
Topic
- April 1, 2025
For the third year running, occupational therapy students have built free assistive devices for local children with disabilities.
Topic
- March 25, 2025
On March 21 at exactly noon, 136 medical students at VP&S opened the envelopes containing their match letters, and learned where they will continue their medical training after graduation.
Topic
- March 26, 2025
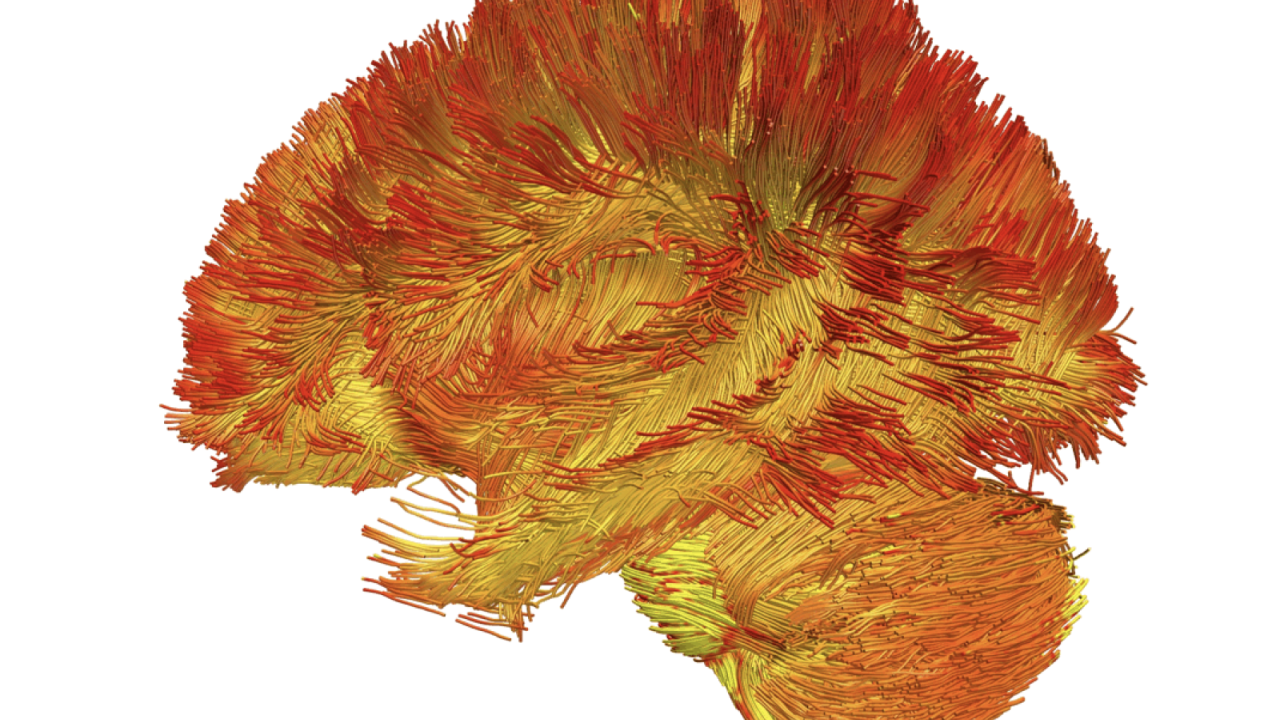
The first atlas of mitochondria in the brain could help scientists develop brain scans that determine the health of a patient’s mitochondria and if they are contributing to disease.
Topic



Our Community

Learn more about our role in serving the Northern Manhattan community of Washington Heights, Inwood, and Harlem.
Diversity, Equity, and Inclusion

At CUIMC, we are committed to providing culturally inclusive medical education, research, and clinical care.